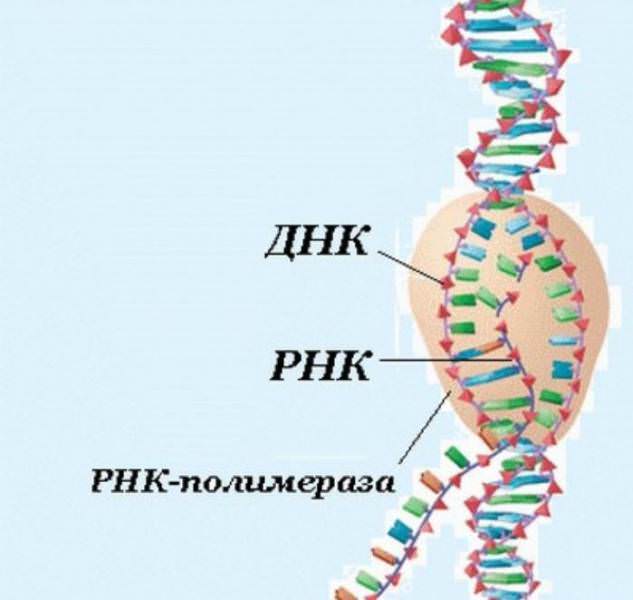
Центральна догма молекулярної біології передбачає, що ДНК містить інформацію для кодування всіх наших білків і три різних типи РНК досить пасивно перетворять цей код у поліпептиди. Зокрема, месенджер РНК (мРНК) переносить білковий план від ДНК клітини до її рибосом, які є «машинами», що керують синтезом білка. Потім РНК (тРНК) переносить відповідні амінокислоти в рибосому для включення в новий білок. Між тим, самі рибосоми складаються в основному з молекул рибосомальною РНК (рРНК).
Однак за півстоліття, що минули з тих пір, як структура ДНК була вперше розроблена, вчені дізналися, що РНК грає набагато більшу роль, ніж просто участь у синтезі білка. Наприклад, було виявлено, що багато типів РНК є каталітичними, тобто вони проводять біохімічні реакції так само, як і ферменти. Крім того, було встановлено, що багато інші різновиди РНК виконують складні регуляторні ролі в клітинах.
Таким чином, молекули РНК відіграють численні ролі як у нормальних клітинних процесах, так і в хворобливих станах. Зазвичай ті молекули РНК, які не приймають форму мРНК, називають некодирующими, оскільки вони не кодують білки. Участь некодирующих мРНК — у багатьох регуляторних процесах. Їх поширеність і різноманітність функцій привели до гіпотези про те, що «світ РНК» міг передувати еволюції ДНК і функцій РНК в клітині, участі у біосинтезі білка.
Некодирующие РНК у еукаріотів
У еукаріотів некодирующая РНК буває декількох різновидів. Найбільш помітно вони переносять РНК (тРНК) і рибосомную РНК (рРНК). Як згадувалося раніше, як тРНК, так і рРНК грають важливу роль в трансляції мРНК в білки. Наприклад, Френсіс Крик припустив існування молекул адапторной РНК, які могли зв’язуватися з нуклеотидним кодом мРНК, полегшуючи тим самим перенесення амінокислот в зростаючі поліпептидні ланцюги.
Робота Hoagland et al. (1958) дійсно підтвердила, що певна фракція клітинної РНК була ковалентно пов’язана з амінокислотами. Пізніше той факт, що рРНК виявилася структурним компонентом рибосом, дозволив припустити: як і тРНК, рРНК також не кодує.
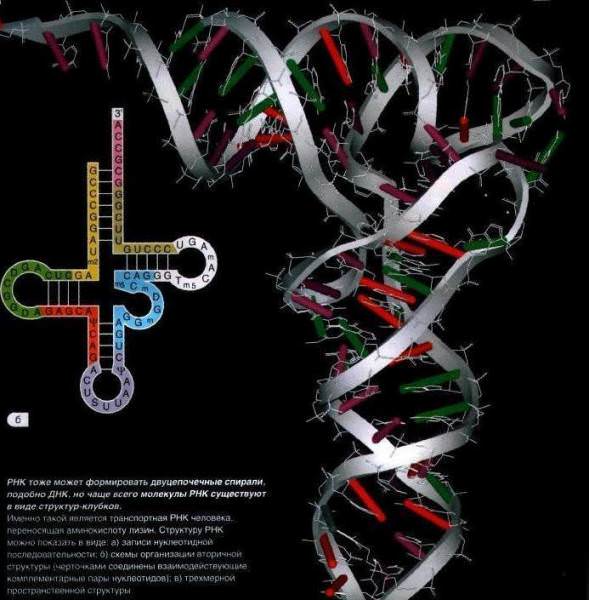
Крім рРНК і тРНК, в эукариотических клітинах існує ряд інших некодирующих РНК. Ці молекули допомагають у багатьох важливих енергетичних, запасаючих функції РНК у клітині, які все ще перераховуються і визначаються. Ці РНК часто називають невеликими регуляторними РНК (сРНК), а в еукаріотів вони були додатково класифіковані по ряду підкатегорій. Разом регуляторні РНК проявляють свої ефекти через поєднання комплементарного спаровування підстав, комплексоутворення з білками і їх власної ферментативної активності.
Малі ядерні РНК
Одна важлива підкатегорія малих регуляторних РНК складається з молекул, відомих як малі ядерні РНК (snRNAs). Ці молекули відіграють важливу роль у регуляції генів шляхом сплайсингу РНК. SnRNAs виявлені в ядрі і, як правило, тісно пов’язані з білками в комплексах, званих snRNPs (дрібні ядерні рибонуклеопротеины, іноді іменовані «snurps»). Найбільш поширеними з цих молекул є частинки U1, U2, U5 і U4 / U6, які беруть участь у сплайсинге пре-мРНК з утворенням зрілої мРНК.
МикроРНК
Інша тема, що представляє великий інтерес для дослідників, — це микроРНК (микроРНК), які являють собою невеликі регуляторні РНК довжиною приблизно від 22 до 26 нуклеотидів. Існування miRNAs і їх скорочувальних функцій РНК в клітині в регуляції генів були спочатку виявлені у нематоди C. elegans (Lee et al., 1993; Wightman et al., 1993). З часу їх відкриття miRNAs вони були виявлені в багатьох інших видах, включаючи мух, мишей і людей. До теперішнього часу було ідентифіковано кілька сотень микроРНК. Може існувати ще багато (He & Hannon, 2004).
Було показано, що микроРНК інгібують експресію генів шляхом репресії трансляції. Наприклад, miRNAs, кодовані C. elegans, lin-4 і let-7, зв’язуються з 3′-нетранслируемым ділянкою їх мРНК-мішеней, запобігаючи утворенню функціональних білків на певних стадіях розвитку личинок. Досі більшість досліджених микроРНК, мабуть, контролює експресію генів шляхом зв’язування з мРНК-мішенями допомогою недосконалого спарювання підстав і подальшого інгібування трансляції, хоча були відзначені деякі винятки.
Додаткові дослідження показують, що микроРНК також відіграють важливу роль при раку та інших захворюваннях. Наприклад, вид miR-155 збагачений В-клітинами, отриманими з лімфоми Беркітта, і послідовність також корелює з відомою хромосомної транслокацією (обмін ДНК між хромосомами).
Малі інтерферуючі РНК
Малі інтерферуючі РНК (мірнк) являють собою ще один клас РНК. Хоча ці молекули мають довжину тільки від 21 до 25 пар підстав, вони також працюють для придушення експресії генів. Зокрема, одна ланцюг дволанцюжкової молекули мірнк може бути включена в комплекс, званий RISC. Цей РНК-містить комплекс може інгібувати транскрипцію молекули мРНК, яка має послідовність, комплементарну її РНК-компоненту.
Мірнк були вперше визначені за їх участі в РНК-інтерференції (РНКи). Вони могли розвиватися як захисний механізм від дволанцюгових РНК-вірусів. SiRNAs отримані з більш довгих транскриптів в процесі, подібному тому, за допомогою якого відбуваються miRNAs і процесинг РНК обох типів залучає той самий фермент, Dicer. Ці два класи, мабуть, розрізняються за їх механізмів репресії, але були знайдені винятку, в яких siRNAs виявляють поведінку, більш типове для miRNAs, і навпаки (He & Hannon, 2004).
Малі РНК Нуклеолярные
Всередині еукаріотичного ядра ядерце є структурою, в якій відбувається процесингу рРНК і рибосомна складання. Молекули, звані малими ядрышковыми РНК (snoRNAs), були виділені з нуклеолярных екстрактів з-за їх великої кількості в цій структурі. Ці молекули функціонують для обробки молекул рРНК, що часто призводить до метилированию і псевдоуридилированию специфічних нуклеозидів. Модифікації опосередковуються одним з двох класів snoRNAs: C / D-бокс або колекції H / ACA-боксів, які зазвичай припускають додавання метильних груп або ізомеризацію урадина у незрілих молекулах рРНК, відповідно.
Некодирующие РНК у прокаріотів
Однак еукаріоти не загнали ринок в некодирующие РНК зі специфічними регуляторними енергетичними функціями РНК у клітині. Бактерії також мають класом невеликих регуляторних РНК. Бактеріальні рРНК беруть участь у процесах, починаючи від вірулентності і закінчуючи переходом від зростання до стаціонарній фазі, який відбувається, коли бактерія стикається з такою ситуацією, як позбавлення поживних речовин.
Одним з прикладів бактеріальної рРНК є 6S РНК, виявлена в Escherichia coli. Ця молекула була добре охарактеризована, її первісну секвенування відбулося в 1980 році. 6S РНК консервативна у багатьох видів бактерій, що вказує на її важливу роль у регуляції генів.
Було показано, що РНК впливає на активність РНК-полімерази (РНКП), молекули, яка транскрибирует РНК-месенджер з ДНК. 6S РНК інгібує цю активність, зв’язуючись з субодиницею полімерази, яка стимулює транскрипцію під час росту. Завдяки цьому механізму 6S РНК інгібує експресію генів, які стимулюють активне зростання, і допомагає клітинам увійти в стаціонарну фазу (Jabri, 2005).
Riboswitches
На регуляцію генів — як у прокаріотів, так і у еукаріотів — впливають регуляторні елементи РНК, звані рибосвитчи (або перемикачі РНК). Рибосвитчи — це РНК-сенсори, які виявляють і реагують на сигнали навколишнього середовища або метаболізму і, відповідно, впливають на експресію генів.
Простим прикладом цієї групи є термодатчик РНК, виявлений в гени вірулентності бактеріального патогена Listeria monocytogenes. Коли ця бактерія проникає у господаря, підвищена температура всередині тіла господаря розплавляє вторинну структуру сегмента в 5′-нетрансліруємій області мРНК, що продукується бактеріальним геном prfA. В результаті цього відбуваються зміни у вторинній структурі.
Було показано, що додаткові рибосвитчи реагують на теплові і холодні удари у різних організмів, а також регулюють синтез метаболітів, таких як цукру, амінокислоти. Хоча рибосвитчи, мабуть, більш поширені у прокаріотів, багато з них також були виявлені в эукариотических клітинах.